|
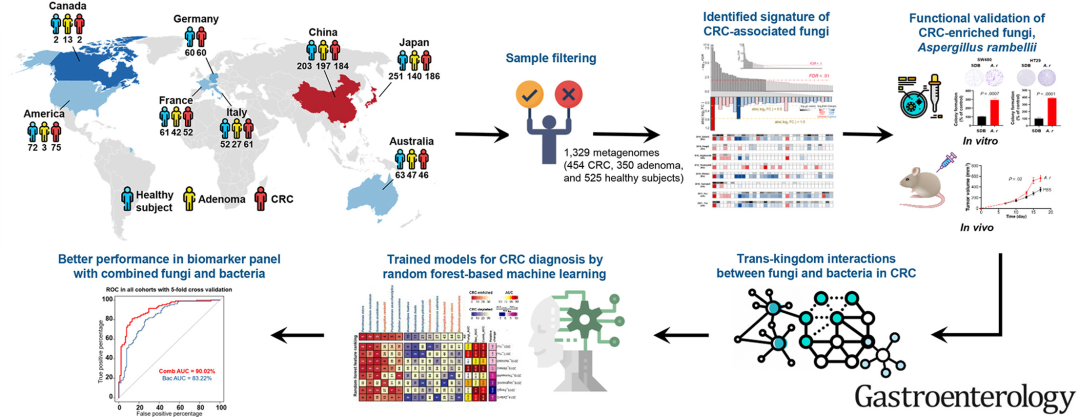
科研| 港中大于君团队揭示结直肠癌的肠道菌群特征和病原真菌(IF:33.883)编译:微科盟小木,编辑:微科盟小编、江舜尧。 导读 背景和目的:肠道菌群是人体肠道微生物群的重要组成部分,但其在结直肠癌(CRC)中的作用仍不明确。我们进行了一项荟萃分析,以揭示真菌菌群对CRC的贡献。 方法:本研究从之前发表的7篇文章中检索了粪便宏基因组数据集,并建立了一个额外的内部队列,共计1329个宏基因组(454个CRC、350个腺瘤和525个健康受试者)。分析了真菌菌群组成和微生物相互作用。候选CRC富集真菌物种(Aspergillus rambellii)在体外和体内进行了功能验证。 结果:多队列分析显示CRC患者肠道菌群发生了改变。我们从多个队列中确定了与CRC和腺瘤患者相关的真菌。标志性CRC相关真菌包括6种富集真菌(Aspergillus rambellii、Cordyceps sp. RAO-2017、Erysiphe pulchra、Moniliophthora perniciosa、Sphaerulina musiva和Phytophthora capsici)和1种减少物种(Aspergillus kawachii)。与腺瘤和健康受试者相比,CRC富集真菌之间的共现相互作用在CRC中变得更强。此外,我们报道了CRC进展过程中肠道真菌和细菌之间的跨界相互作用,其中A. rambellii与CRC富集细菌具核梭杆菌(Fusobacterium nucleatum)密切相关。A. rambellii促进体外CRC细胞生长和异种移植小鼠的肿瘤生长。我们进一步发现,真菌和细菌联合生物标志物比纯细菌组更准确地区分CRC患者和健康受试者(AUC相对变化增加1.44%-10.60%)。 结论:本研究揭示了结直肠肿瘤发生阶段的肠道菌群特征和病原真菌。除细菌外,粪便真菌还可用于CRC患者的无创诊断。 图文摘要 论文ID 原名:Altered mycobiota signatures and enriched pathogenic Aspergillus rambellii are associated with colorectal cancer based on multi-cohort fecal metagenomic analyses 译名:基于多队列粪便宏基因组分析,改变的真菌菌群特征和富集的致病性Aspergillus rambellii与结直肠癌相关 期刊:Gastroenterology IF:33.883 发表时间:2022.6.18 通讯作者:于君 通讯作者单位:香港中文大学 DOI号:10.1053/j.gastro.2022.06.038 实验设计 结果 1 共有1329名受试者被纳入CRC进展的肠道菌群荟萃分析 在这项荟萃分析中,我们纳入了8项已发表的研究,这些研究使用鸟枪法宏基因组学来表征CRC患者与健康受试者的粪便微生物群(表S1和S2)。我们还建立了一个内部粪便宏基因组数据作为额外的研究群体(表S1和S2)。在这些宏基因组中,中位数为0.16%(10 -2.80 )的总宏基因组reads被映射到真菌基因组(图1A和S表2),这与之前的研究一致(0.1%)。稀疏曲线显示,所有队列样本在真菌测序reads达到10,000时均达到平台状态(图1B)。为了确保样品质量,我们采用了一系列严格标准的过滤来去除污染或真菌含量低的样品(图1C)。Hanningan等人的队列进行了PCR预处理,并且测序深度较低,因此该数据集被丢弃以避免后续分析中的扩增偏差。过滤后,共保留了1329个宏基因组,包括454名健康受试者(占检索到的健康宏基因组总数的68%)、350名腺瘤患者(占检索到的腺瘤宏基因组总数的73%)和525名CRC患者(占检索到的CRC宏基因组总数的68%)(图1D)。CRC、腺瘤和健康受试者的丢弃样本没有显著差异,表明我们的样本均匀分布在不同的研究对象组中。 图1 宏基因组测序数据及病例筛选标准。(A)左图:9个队列(n = 2052)中真菌(红色边框)或细菌(蓝色边框)排列计数。右图:所有样本中真菌与细菌排列计数的比率(紫色边框)。(B)基于真菌chao1多样性生成的微生物群稀疏曲线。(C)病例过滤标准包括3个主要部分:样本序列质量控制、疑似污染样本过滤和低真菌序列深度样本丢弃。总共有723例被丢弃,与之前的研究一致,大约30%的受试者没有真菌。(D)选择的(黄色)和丢弃的(深蓝色)样本在检索到的CRC、腺瘤和健康受试者宏基因组中所占的百分比。丢弃样本均匀分布在不同的研究对象组中(χ2 =4.03,p = 0.13;卡方检验)。 2 肠道菌群在CRC进展中发生改变 首先研究了门水平的真菌组成。在所有队列中,子囊菌门(Ascomycota)是CRC患者和健康受试者中最普遍的真菌门(图S1A和S1B)。然后,我们比较了CRC患者和健康受试者之间前5个门的丰度,发现这些门在所有队列中都没有一致的丰度变化(图S1C)。对于真菌丰富度,我们发现将所有队列组合在一起时,CRC患者的α多样性(chao1指数)显著降低,这与之前的报告一致( p <0.05)(图S1D)。而仅在三个队列中观察到α多样性的显著差异,表明存在队列间变异。使用主坐标分析(PCoA),我们发现各队列间真菌群落的变化显著归因于人群(洲或性别)和疾病阶段(CRC、腺瘤或健康)的差异(图S2),与之前的研究一致。体重指数(BMI)和肿瘤部位对真菌群落变化没有显著影响。总体而言,我们证实了肠道真菌菌群与CRC的关联。 3 确定CRC和腺瘤中的标志性真菌种类 为了表征CRC相关真菌,将所有CRC患者中相对丰度>0.1%(n=296)的单个真菌丰度与健康受试者进行了比较(表S3)。在FDR<0.1时,我们确定了74种真菌物种,其丰度存在差异,然后在FDR<0.01时,我们将其缩减为入围的33种(下文称为真菌核心组)(Wilcoxon秩和检验)(图2A和S表7)。该核心组由与CRC相关的具有不同丰度的物种组成,其中 Aspergillus rambellii 是最丰富的,而 Aspergillus kawachii 最少(图2B)。每个真菌物种的丰度变化在队列间差异很大,因此表明存在队列间差异(图2C)。我们进一步评估了队列异质性和潜在混杂因素对单个真菌物种的影响。与我们的PCoA分析(图S2)一致,队列异质性比其他因素(年龄、BMI和性别)的影响更大(图S3)。值得注意的是,与队列异质性和其他混杂因素相比,疾病阶段(CRC、腺瘤或健康)与33个物种的真菌核心组丰度的变化更密切相关,表明疾病阶段对物种组成的影响大于队列间变异。 真菌核心组包括10种CRC富集真菌和23种CRC减少真菌(图3A)。与健康受试者相比,CRC患者的 A. rambelli 丰度增加3.0倍, A. kawachii 丰度减少2.0倍( p <0.0001)(图3B)。与健康受试者相比,CRC患者中 A. rambelli 的中位丰度更高,并且这种增加在8个队列中的7个队列中持续显著(图3C;表S8和S9)。为了确定在不同队列中一致变化的真菌,我们识别了8个队列中至少在7个队列中发生变化的18种真菌(7种CRC富集,11种CRC减少)(表S9)。7种一致的CRC富集真菌分别是 A. rambellii、Erysiphe pulchra、Aspergillus ochraceoroseus、Elsinoe australis、M. perniciosa、Sphaerulina musiva 和 Thielaviopsis punctulata 。11种一致的CRC减少真菌分别为 Trichophyton mentagrophytes 、 olletotrichum fioriniae 、 Colletotrichum salicis 、 Colletotrichum tofieldiae 、 Paracoccidioides brasiliensis 、 Pichia kudriavzevii 、 Pneumocystis murina 、 Pseudocercospora fijiensis 、 Rhizoclosmatium globosum 、 Rhizoctonia solani 和 Spizellomyces punctatus 。此外,在真菌核心组中,从CRC前体、早期CRC到晚期CRC,发现了2种真菌( Cordyceps sp. RAO-2017 和 Phytopythium vexans )显著且一致的富集,1种真菌( A. kawachii )减少,表明这些真菌物种在CRC进展中的因果作用(图S4)。 然后将CRC患者与腺瘤患者的真菌丰度进行比较。FDR<0.01时,在CRC和腺瘤患者之间发现了24种差异丰度真菌(图S5A和表S10)。在这24种真菌中,观察到1种CRC富集( A. rambellii )和6种CRC减少的真菌物种( A. kawachii、Fusarium pseudograminearum、Lentinula edodes、Hanseniaspora guilliermondii、Saitoella complicata 和 Cryptococcus neoformans )的丰度变化是腺瘤患者的1.4倍以上(S图5B)。值得注意的是,当将CRC患者与腺瘤患者和健康受试者进行比较时,7种真菌的丰度发生了一致的改变(表S7和10),这表明它们是CRC腺瘤相关真菌的标志性物种。这7种真菌包括6种CRC富集真菌( A. rambellii、Cordyceps sp. RAO-2017、E.pulchra、Moniliophora perniciosa、S. musiva 和 Phytophthora capsici )和1种CRC减少真菌( A. kawachii ) (图S6)。 我们进一步比较了腺瘤患者和健康受试者之间的真菌丰度。FDP<0.01时,在腺瘤患者和健康受试者之间发现了10种差异丰度真菌(图S7A)。在这10种真菌中,腺瘤患者中5种真菌的丰度至少增加了1.5倍(图S7B)。此外,与CRC患者和健康受试者相比,腺瘤患者中的5种真菌(包括 Cryptococcus neoformans 、 Encephalitozoon cuniculi 、 F. pseudograminearum、S. complicata 和 Talaromyces islandicus )发生了显著改变(表S10和S11)。当CRC患者与健康受试者比较时,这5种真菌未被纳入核心组(表S7和S11),表明这5种真菌仅与腺瘤相关,而与CRC无关。总之,我们的多队列宏基因组分析揭示了与CRC和腺瘤相关的标志性真菌物种。 图2 多队列分析报告了一组与CRC密切相关的肠道真菌。(A)相对丰度<0.1%的296种真菌中,发现了CRC患者和健康受试者之间丰度显著变化的真菌(n=74,FDR<0.1;n=33,FDR<0.01)。(B)在所有队列中出现成对倍数变化的真菌核心组(n=33)。红色和蓝色条分别代表CRC富集和减少的真菌物种。倍数变化大于两倍或小于一半的真菌被突出显示。(C)每个队列中具有成对倍数变化的真菌核心组。采用双侧Wilcoxon检验,并在每个单独研究中概括倍数变化。Abs,绝对值;FC,倍数变化。 图3 CRC患者与健康受试者的特征真菌种类比较。(A)真菌核心组包括10种CRC富集真菌和23种CRC减少真菌物种,其在CRC患者(n=454)、腺瘤患者(n=350)和健康受试者(n=525)之间的丰度存在显著差异。真菌丰度归一化为相对于对照样品丰度中值的自然对数倍数变化。临床和队列信息显示在图顶部,包括队列、性别、年龄、肿瘤位置和BMI。(B, C)所有队列(B)和每个研究(C)中倍数变化最大的两种真菌A. rambellii(CRC富集)和A. kawachii(CRC减少)的小提琴图。健康受试者和CRC患者分别用黄色和红色表示。 4 Aspergillus rambellii在体外和异种移植小鼠模型中促进CRC 为了验证计算机实验结果,将两种CRC细胞系SW480和HT29以及一种正常结肠上皮细胞系NCM460与 A. rambellii ( A.r CM)条件培养基或对照培养基SDB共培养。 A.r CM显著增加了CRC和正常上皮细胞的活力(SW480和NCM460, p < 0.01;HT29, p < 0.001)(图4A)。 A.r CM还显著促进了肿瘤细胞集落的形成(SW480和HT29, p < 0.001)(图4B)。然后,我们将两种源自CRC患者的类器官与 A.r CM或SDB对照共培养。 A.r CM处理后,CRC类器官的生长和增殖得到了显著促进(图4C)。然后,我们使用植入CRC细胞系SW480的皮下异种移植小鼠模型来评估 A. rambellii 在体内是否具有致癌作用。瘤内注射 A. rambellii 或PBS作为对照,我们发现与PBS对照相比, A. rambellii 显著促进肿瘤生长( p < 0.05)(图4D)和肿瘤重量( p < 0.05)(图4E)。与对照小鼠的肿瘤组织相比, A. rambellii 还显著促进了肿瘤组织中的细胞增殖,表现为Ki-67阳性细胞比例较高( p < 0.001)(图4F)和PCNA蛋白水平升高( p < 0.0001)(图4G)。因此,我们在体外和体内研究的一致结果表明, A. rambellii 可以促进CRC进展。 图4 Aspergillus rambellii在体外和体内促进结直肠肿瘤的发生。(A) A.r CM(10.0%)促进CRC细胞(SW480, HT29)和正常结肠上皮细胞(NCM460)的活力。以SDB培养基为对照。(B) A.r CM增强了CRC细胞集落的形成。(C)来自CRC患者的类器官(828和816)用A.r CM治疗。左图:A.r CM或SDB处理的类器官培养6天后的代表性图像(比例尺=100 μm)。右图:类器官表面积(104 μm2)。(D-G) SW480异种移植小鼠分别用A. rambellii处理或PBS对照。A. rambellii显著促进肿瘤生长(D)、肿瘤大小和肿瘤重量(E)。(F) Ki67阳性细胞的代表性免疫组化图像及定量数据分析。放大倍数200×,比例尺=30μm。(G) SW480异种移植物中PCNA蛋白表达水平,用β-actin表达标准化。数据用平均值±标准差表示。A.r和PBS处理分别用红色和黑色表示。 5 CRC进展中的真菌-真菌和真菌-细菌相互作用 已知微生物之间的相互作用与结直肠肿瘤的发生有关。因此,我们通过与DGCA的相关性分析,评估了CRC、腺瘤和健康受试者中33种核心真菌物种之间的相互作用。真菌之间的共发生和共排斥相互作用在不同的受试者组之间存在显著差异,这种真菌-真菌相互作用对CRC的影响逐渐增强(图5A和图S8)。在CRC中,4种CRC富集真菌(包括 A. rambellii 、 E. pulchra 、 T. punctulata 和 S. musiva )显示出显著的共现中心性。而这些共现相关性在腺瘤患者中较弱,在健康受试者中则消失。与CRC相比, A. rambellii 与腺瘤和健康受试者中的其他真菌具有完全不同的相互作用,仅与 M. perniciosa 有很强的相关性。我们的研究结果确定了关键真菌(特别是 A. rambellii ),它们在CRC进展过程中的肠道菌群中具有核心作用。 鉴于人类肠道微生物群以细菌为主,我们评估了真菌和细菌之间的相互作用。与健康受试者相比,在CRC中鉴定出31种差异丰度细菌(以下称为细菌核心组),包括致病性具核梭杆菌( Fusobacterium nucleatum ) 、Parvimonas micra 和 Gemella morbillorum 的富集,以及益生菌 Roseburia intestinalis、Bifidobacterium bifidum 和 Streptococcus thermophilus 的减少(图S9和表S12)。然后,我们评估了真菌和细菌之间的跨界相互作用,观察到从健康受试者(n=143)、腺瘤患者(n=156)到CRC患者(n=184),真菌-细菌的显著相互作用数量有所增加(图S10和表S13)。同样,真菌-细菌相互作用的强度也随着CRC的进展而增强(图5A和图S10)。共有99种真菌-细菌相互作用,包括 A. rambellii 和 F. nucleatum 之间的相关性,仅在CRC中显著,而在腺瘤和健康受试者中不显著(图5B)。而在CRC和腺瘤中,共有17种真菌-细菌相互作用(包括 A. rambellii 和 P. micra 之间的相关性)均显著。总体而言,我们揭示了CRC进展中肠道真菌和细菌之间的跨界相互作用。 图5 CRC进展中的真菌-真菌和真菌-细菌相互作用。(A)左图:CRC、腺瘤和健康受试者中33种真菌核心组和31种细菌核心组之间的相关性。右图:不同疾病阶段的33种真菌核心组之间的相关性。粉色菱形、蓝色圆圈和绿色圆圈分别代表CRC与健康受试者、CRC与腺瘤、腺瘤与健康受试者之间显著不同的配对相关性。(B)热图显示跨界真菌-细菌相关性与疾病进展的一致增加或减少趋势。仅显示一致的相关性。 6 肠道真菌和细菌之间的相互作用与CRC进展相关 通过比较CRC与健康受试者的相关性强度,进一步研究了微生物相互作用是否与CRC进展相关。我们发现真菌-真菌相互作用网络(z-score=+4)不同于细菌-细菌相互作用网络(z-score=-2)(图S11A)。然后定义了CRC与健康受试者之间的9类差异配对相关性(S图11B)。在健康受试者中观察到更多正的真菌-真菌差异相关性(占总真菌-真菌差异相关性的30.77%),随着CRC的进展而下降。相比之下,CRC中大多数真菌-细菌差异相关性为正(占总真菌-细菌差异相关性的56.72%),但在健康受试者中不显著。 为了可视化差异相互作用网络,使用DGCA并通过相似性传播簇将具有相似差异相关强度(z-score)的微生物分组在一起。33种真菌和31种细菌的核心组被分成6个簇(图6),这6个簇中有2个表现出优势,分别被命名为Fun_cluster(81.8%的微生物是真菌)和Bac_cluster(81.0%的微生物是细菌),而这两个大集群也暗示了CRC进展中真菌-真菌和细菌-细菌相互作用之间的差异。值得注意的是,Fun_cluster中包含了3种成熟的CRC富集细菌,即 F. nucleatum 、 F. periodonticum 和 P. micra ,并显示出与其他真菌的强差异相关性。例如, A. rambellii 与 F. nucleatum 和 P. micra 的相关z值分别为-5.95和-5.07(表S13),表明致病菌可能与CRC中的真菌有密切的相互作用。对于Bac_cluster,涉及的大多数微生物是益生菌,如 S. thermophilus 和 Eubacterium eligens 。总之,我们的差异相关分析揭示了CRC进展中真菌和细菌之间的相互作用。 图6 CRC与健康受试者之间的差异相关网络。采用差异相关指数(z-score)评估CRC与健康受试者之间微生物相互作用的差异。通过相似性传播簇将6个集群聚在一起。这两个主要集群被称为Fun_cluster和Bac_cluster,因为它们分别主要包括真菌和细菌。Abs(z-score),z-score绝对值、CorrCRC,CRC患者的相关性;CorrCTRL,健康受试者的相关性;Countscorr、相关数量。 7 结合真菌和细菌的生物标志物组可提高CRC诊断性能 因此,我们评估了肠道真菌是否也可以作为CRC的诊断生物标志物。在33种真菌和31种细菌的核心组中,只有4种在区分所有8个队列中的CRC患者和健康受试者时的中位AUC大于60%,包括 A. rambellii (图S12A)和3种细菌( F. nucleatum、P. micra 和 G. morbillorum )(图S12B)。 P. micra 的诊断准确性最高,中位AUC为67.95%,但这种表现在临床上被认为不令人满意。为了提高准确性,我们采用了一种结合了真菌和细菌的生物标志物组。我们用纯真菌(n=17)、纯细菌(n=12)或两者混合(5种真菌和9种细菌)对CRC诊断小组进行培训(图7A和图S13)。在两个队列(2016_VogtmannE和2019_WirbelJ)中,纯真菌组诊断CRC的性能高于纯细菌组。然后,我们将来自8个队列的所有样本合并在一起,通过随机森林和5倍交叉验证,评估生物标志物组的整体性能,以区分CRC患者和健康受试者(图7B)。真菌与细菌联合组的AUC达到90.02%,纯细菌组的AUC为83.22%。与纯细菌组相比,真菌和细菌联合组的性能得到了很大提高,8个队列中有5个队列的AUC达到80%以上(AUC提高了1.44%-10.60%)(图7C)。真菌和细菌联合组也可以将早期CRC(AUC=73.85%-89.85%)或晚期CRC患者(AUC=80.76%-90.86%)与健康受试者区分开来(S图14)。此外,与纯细菌组(AUC=82.95%)相比,联合组可以将CRC与腺瘤患者进行分类(AUC=86.39%),性能更好(图S15)。因此,我们的研究结果表明使用真菌和细菌混合标志物的诊断小组比使用纯细菌的常规小组更准确地识别CRC患者和健康受试者。还测试了我们的生物标志物组在腺瘤患者中的诊断性能。我们训练了分类器以使用相同的方法从健康受试者中识别腺瘤患者。纯真菌组(n=11)用于鉴别腺瘤患者的平均AUC为57.17%(图S16A),将该组与细菌生物标志物相结合可提高其性能(AUC=68.44%;6种真菌和4细菌)(图S16B-D)。 图7 结合真菌和细菌物种的生物标志物组提高了CRC诊断性能。(A)上图:使用内部随机森林得分估计的每个队列中每个微生物物种对交叉验证预测性能的重要性(橙色:真菌;蓝色:细菌)。仅包括出现在至少一个数据集中的三个顶级特征中的物种。下图:8个队列中真菌、细菌(Bac)和真菌-细菌联合(Comb) CRC诊断模型的AUC表现,以及细菌和真菌-细菌联合诊断模型之间的相对变化。(B, C)通过随机森林对所有样本进行5倍交叉验证(B),或在每个队列中使用LOSO(C),将CRC患者与细菌(Bac)和真菌-细菌(Comb)分类器的健康受试者区分开来的诊断性能。 讨论 在本研究中,我们检索了之前7项CRC微生物群研究的宏基因组数据集,并建立了一个额外的内部队列来揭示CRC中真菌菌群的分布情况。细菌与CRC的关联已得到广泛认可,而真菌由于其在肠道微生物群中的丰度相对较低而经常被忽视。这种低丰度极大地影响了对这些真菌的研究,因为单个宏基因组数据集可能没有足够的测序reads和样本量来获得肠道菌群的全面概况。同时,通过结合可比研究的结果,多队列分析可以提高统计能力,减少队列特异性偏差。在本研究中,我们揭示了来自不同人群的CRC患者肠道真菌菌群的改变,我们的结果清楚地表明,CRC进展过程中(从健康受试者、腺瘤患者到CRC患者)真菌生态失调的发生。 然后,我们确定了与CRC不同进展阶段相关的标志性真菌物种。特别是,从不同人群中确定了7种与CRC患者一致相关的特征真菌,包括6种富集真菌( A. rambellii 、 Cordyceps sp. RAO-2017 、 E. pulchra、M. perniciosa、S. musiva 和 P. capsica )和1种减少真菌( A. kawachii )。一些与CRC相关的真菌,如 Aspergillus flavus 、 Kwoniella mangrovensis 和 Pseudogymnoascus sp. VKM F-4518 在我们之前的单一人群研究中被报道过。不同地域人群的肠道微生物群差异很大。事实上,我们发现CRC相关的真菌在人群中是多样化的。为了识别普遍的CRC相关真菌,我们选择了在多个人群中不断改变的物种(如 A. rambellii )。值得注意的是,队列间变异是微生物荟萃分析中的一种常见现象,因为不同的研究有不同的样本收集和测序技术方法,而来自不同地理区域的个体肠道微生物群的高度动态特性可能进一步导致生物变异。因此,我们研究了队列异质性和潜在混杂因素(如年龄、性别、BMI和肿瘤位置)对真菌群落的影响。尽管存在队列间差异,但我们发现疾病阶段(CRC、腺瘤或健康)对物种组成的影响最大(图S3),进一步支持了我们发现的普遍的CRC相关真菌独立于人群差异。 我们的多队列宏基因组分析共同揭示了整个真菌群落随着健康受试者-腺瘤-CRC序列中真菌丰度的显著变化而改变。在我们的33种真菌核心组中,我们发现 A. rambellii 在健康受试者、腺瘤和CRC序列中一致富集, A. kawachii 一致减少(图3)。为了证实已鉴定的CRC相关真菌的作用,我们进行了功能实验,以验证 A. rambellii 的致癌特征,这是CRC患者中最丰富的物种。我们发现 A. rambellii 显著促进体外癌细胞和体内肿瘤的生长,从而证实了真菌失调在CRC中的潜在因果关系。这些发现得到了先前研究的支持,这些研究表明 A. rambellii 能够产生多种黄曲霉毒素(如黄曲霉毒素B、黄曲霉毒素G)和黄曲霉毒素前体杂色曲霉素。黄曲霉毒素是真菌衍生的毒素,已被归类为致癌物和诱变剂 ,其促进肝癌发生的强大能力就是例证。例如,伴有乙型肝炎感染的AFB1长期饮食暴露与肝癌风险显著增加有关。AFB1因其对免疫系统的毒性作用而闻名,因为它可以通过激活toll样受体损害人体巨噬细胞和树突状细胞,从而诱导免疫失调。虽然曲霉属( Aspergillus )的其他真菌,尤其是 A. flavus 与癌症的关联经常被报道,但在这里我们首次揭示了 A. rambellii 对促进CRC进展至关重要。迄今为止,关于AFB1与CRC相关性的研究很少,因此需要进一步的机制研究来评估 A. rambellii 是否产生AFB1或其他黄曲霉毒素以促进结直肠肿瘤的发生。有趣的是,我们发现另一种曲霉属真菌 A. kwachii 在CRC患者中显著减少。这表明同一属的不同物种可能产生不同甚至相反的效应,从而强调了在物种水平上进行微生物群研究的重要性。此外, C. neoformans 、 E. cuniculi 、 F. pseudograminearum 、 S. complicate 和 T. islandicus 是与腺瘤患者相关的5种标志性真菌。 众所周知,微生物在人体肠道微生物群中共存,而它们之间的相互作用可能与健康和疾病有关。例如,我们之前的研究报告称,CRC中共存细菌相关性变得更加普遍。本研究揭示了CRC或腺瘤患者的真菌相关性网络存在显著差异。在CRC的相关网络中,4种CRC富集真菌(包括 A. rambellii 、 E. pulchra 、 T. punctulata 和 S. musiva )显示出共现中心性。与腺瘤患者和健康受试者相比,CRC中 A. rambellii 和其他真菌之间的相关性发生了很大变化。值得注意的是,我们发现 CRC中整体真菌-真菌相关性降低,而细菌-细菌相关性在CRC进展过程中增强。 这些结果表明,肠道真菌和细菌在CRC中可能存在拮抗作用:健康个体中的真菌相互作用可能会因结直肠肿瘤发生过程中细菌组成的改变而被破坏。细菌-细菌相关性的增加可能会影响肠道真菌群落,从而为致病菌的生长提供有利条件。 然后,我们评估了CRC进展中肠道真菌和细菌之间的跨界相互作用。我们的结果显示,CRC中大多数真菌-细菌相关性为正相关,但在健康受试者中不显著。而一些真菌物种与致病菌密切相关,例如 A. rambellii 与公认的CRC相关细菌 F. nucleatum 显著正相关。以往的研究一致报道了几种真菌和细菌物种在CRC进展中的共现相关性。人体肠道微生物群由不同界的微生物组成,包括细菌、真菌和病毒,因此可以合理地认为,跨界微生物相互作用在健康和疾病条件下经常发生。我们之前报道了CRC中肠道细菌和病毒之间的跨界相互作用,而在本研究中我们的分析表明真菌和细菌之间的相互作用参与并促进了CRC的进展。值得注意的是,这些发现仅基于宏基因组测序分析,因此需要进行机理研究来验证结直肠肿瘤发生中的微生物相互作用。 大量研究显示了使用细菌生物标志物诊断CRC的潜力。本研究证明了粪便样本中的真菌种类也可以作为生物标志物来识别8个队列中的CRC患者,平均AUC为73%。而 真菌和细菌混合的生物标志物组在诊断性能上有很大的提高 ,平均AUC为83%。与纯细菌生物标志物组相比,8个队列中有7个队列的AUC相对变化增加了1.44%-10.60%。测量粪便中的微生物丰度由于其侵袭性低于传统的结直肠恶性肿瘤内镜筛查,因此在世界范围内得到越来越多的认可。然而,在群体水平上识别特征微生物一直具有挑战性,因为不同人群的肠道微生物群具有高度异质性。此前,Wirbel等人和Thomas等人的荟萃分析报道了几种CRC富集细菌物种及其作为CRC诊断生物标志物的能力。在本研究中,我们的多队列分析表明,真菌和细菌生物标志物的联合使用可以进一步提高诊断性能,从而凸显了肠道真菌在临床应用中的潜力。 结论 综上,我们的荟萃分析阐明了肠道真菌菌群在CRC进展中的重要作用。我们确定了与来自不同人群的CRC和腺瘤患者一致相关的标志性真菌,并揭示了致病性 A. rambellii 在体外和体内显著促进了CRC的发展。因此,有必要进行进一步的机制研究来表征菌群失调的后果,并评估CRC富集真菌物种是否是CRC的原因。我们的研究结果还强调了真菌作为CRC诊断生物标志物的潜力。有必要对结合真菌和细菌生物标志物的CRC诊断组进行临床评估,这可能会改善当前基于单一细菌界的生物标志物组的性能。 原文链接: https://www.sciencedirect.com/science/article/pii/S0016508522006564 |